بسم الله الرحمن الرحيم,
من بين التصورات التي ترسبت لدينا من المقررات الدراسية و حتى الجامعية لدى تخيلنا لعلمية النسخ هو أن للحمض النووي بنية خطية تلتصق بها بروتينات و أنزيمات مختلفة كالأرنا بوليميراز II ومجموعة من عوامل النسخ لتصنيع الـ RNA . لكن إذا أخذنا في عين الاعتبار العلاقة بين طول الحمض النووي و حجم النواة للاحظنا أنه في الطور الاستوائي (Metaphase) يتم ضغط الحمض النووي حوالي عشرة آلاف مرة, مما يعطينا فكرة أن للحمض النووي بنية ثلاثية الأبعاد. يتم ضغط الحمض النووي بواسطة بروتينات هستونية و غير هستونية لتشكيل ما يسمى بـ "الكروماتين" . الأمر المذهل في هذا هو أن طبيعة هذا الضغط لا تمنع من أن يكون للحمض النووي بنية مرنة تتيح لعوامل مختلفة الوصول إلى عدة مناطق من أجل تعديل مختلف الوظائف الخلوية كعملية النسخ, تصليح الحمض النووي و تعديل التعبير الجيني.
يدل التباين في نشاط الجينات والوظائف الخلوية في مختلف العوامل الزمنية والبنيوية ومن خلية إلى أخرى على أن للبنية الثلاثية للكروموزموم أهمية بارزة في تعديل النشاط الجيني و أن فهم هذه العلاقة سوف يمكننا من فهم الميكانزمات الخلوية ويساعدنا على تفسير بعض المشاكل في العمليات الخلوية كالسرطان مثلا, إذا أن واحدة من أهم خصائص السرطان هو الاختلال في بنية الكروماتين و سلسلة الحمض النووي.
لحسن الحظ ساهم ظهور آلات السَلْسَلة وتقنيات الترسيب المناعي في تطوير تقنيات جديدة لدراسة هذه البنية وعلاقتها بالتعديل الجيني على نطاق شامل للجينوم. كما ساهمت في المرور من دراسة بنية الكرموزم باستعمال طرق غير كمية وضعيفة الدقة كالميكروسكوب وطريقة FISH لتلوين الحمض النووي إلى استعمال طرق كمية أكثر دقة .
مستويات ضغط الكروماتين
سمح ظهور تقنيات المكروسكوب الضوئية برؤية البنية التشكيلية للكروماتين في مختلف مستويات ضغطه حيث يصل الكروماتين إلى أعلى مستوى ضغط في المرحلة الاستوائية من الانقسام الخلوي على شكل صبيغيات لينفتح بعدها في الحالة الطبيعية للخلية. يتم التحكم في نسبة هذا الانفتاح والانغلاق عن طريق مجموعة من البروتيات الهستونية يلتف حولها الحمض النووي بالإظافة إلى بروتينات أخرى تتحكم في نسبة هذا الالتفاف. لاحظ الباحثون أن طريقة تنظيم الكروماتين ليست بعشوائية وأنها تختلف على حسب الخلية فيتم فتح وغلق مناطق من الجينوم على حسب إحتياج الخلية فمثلا في خلايا الدم يتم فتح المناطق التي تحتوي على الجينات المسؤلة عن تصنيع بروتينات الجلوبين ويتم تثبيطها في الخلايا الأخرى.
يمكن تصنيف مستويات تنظيم بنية الكروماتين إلى أربع مستويات:
1. المستوى الأول (لفائف 11nm): ويعتبر المستوى الأساسي لتنظيم الكروماتين حيث يلتف 147 جزئي قاعدي من الحمض النووي على 8 بروتينات ( نسختان من H2A, H2B, H3, H4 ) لتشكل مايسمى بـ النيكليوزوم ذا قطر بحوالي 11nm. تطلق تسمية " خرزات على الخيط" على تتابع سلسة من النكليوزمات نظرا لشكلها تحت المكروسكوب المشابه لمجموعة خرزات موصولة بحبل. يترواح طول المنطقة الفاصلة بين نكليوزومين من 20 إلى 90 حمض قاعدي على حسب الخلية ومن كائن إلى آخر, تسمى هذه المنطقة بالحمض النووي الرابط (linker DNA).
2. المستوى الثاني (لفائف 30nm) : تؤدي إظافة الهستون H1 أو H5 (تسمى أيضا الهستونات الرابطة, Linker Histone) إلى النكليوزمات إلى تشكيل بنية أكثر ضغطا ذات قطر بحوالي 30nm.
3. المستوى الثالث (عروات لفائف 30mn): إلى غاية المستوى الثاني يبقى الكروماتين 100 مرة أكبر من حجم النواة لهذا يتم تنظيم لفائف الـ 30nm على شكل حلقات. في الطور البنيوي للخلية ( interphase) (الطور الذي تنمو فيه الخلية وتزاول فيه نشاطها) تكون الكروموزمات مقسمة إلى نطاقات متكونة من مجموعة عروات الكروماتين (Chromatin loops) متكونة من لفائف الـ 30mn. تكون أغلبية لفائف الـ 30mn مضغوطة بشكل أكبر لتشكل ما يسمى بالكروماتين المغاير ( heterochromatin) مما يؤدي إلى اسكات الجينات المتواجدة في تلك المناطق. وخلافا لمناطق الكروماتين المغاير توجد مناطق تسمح بدخول آلات النسخ و تتميز بنشاط جيني تسمى الكروماتين الحقيقي (euchromatin).
تختلف مناطق الكروماتين المغاير من خلية إلى أخرى حيث يتم ضغط وفتح مناطق من الحمض النووي على حسب الحاجة اليها, تسمى مناطق الكروماتين المغاير التي يمكن أن تنفتح وتتحول إلى مناطق كروماتين حقيقي بالكروماتين المغاير الاختياري ( facultative heterochromatin) والمناطق التي الثابتة نسبيا (والمتواجدة بقرب السنترومير والتيلومير) بالكروماتين المغاير الأساسي.
4. المستوى الرابع (التوزيع الثلاثي الأبعاد للكروموزوم في النواة): يمثل هذا المستوى التموضع النسبي للكروموزمات بالنسبة لبعضها البعض وبالنسبة للصحيفة النووية ( lamina) للغشاء النووي. في العادة يزداد نشاط الجينات كلما ابتعدت عن الصحيفة النووية.
رسم بياني يبين مختلف مستويات تنظيم الكروماتين (مصدر الصورة)
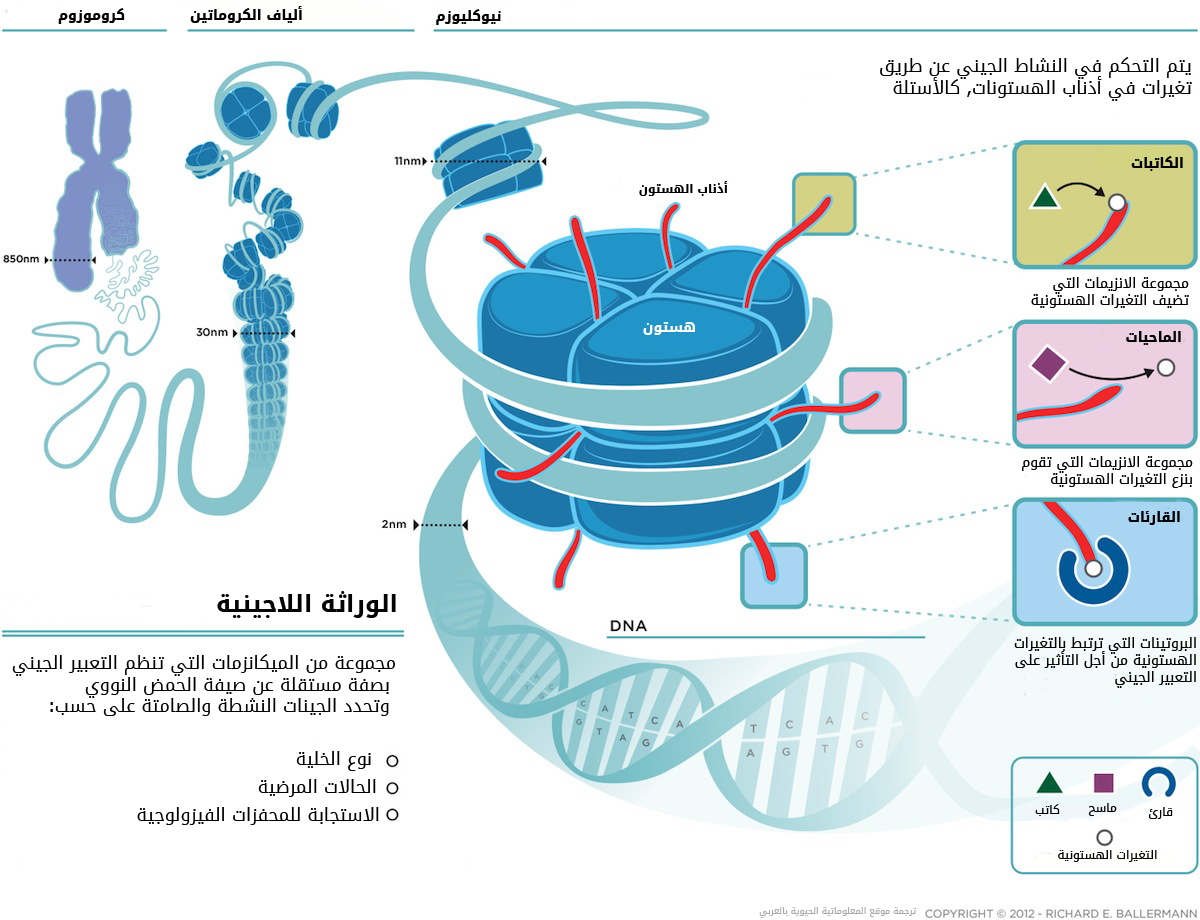
يتم التحكم في بنية الكروماتين عن طريقة تغيرات لاجينية ( epi-genetic modifications) تؤثر في الخصائص الكيمائية للهستونات وبعض قواعد الحمض النووي. تتم التغيرات الهستونية على أذناب البروتينات الهستونية من أجل تغير تكوين الكروماتين و تهيئة مناطق ارتباط معقدات بروتينية و أنزيمية. تصنف هذه الأنزيمات والبروتينات على حسب وضيفتها إلى :
| الكاتبات (writers) | تقوم بوضع التغيرات الهستونية |
| الماحيات ( Erazers) | تقوم بنزع التغيرات الهستونية |
| القارئات (Readers) | البروتينات التي تقوم بفسير التغيرات الهستونية و التأثير على التعبير الجيني |
وضع العلماء طريقة لتسمية التغيرات التي تحدث في أذناب الهستونات حيث يتم كتابة اسم الهستون التي حدتت فيها التغيرات والوضعية التي حدثت فيها التغيرات ونوع التغير, مثلا H3K4me3 تدل على أن التغير حدث في الهستون H3 في الوضعية K4 وان نوع التغير هو ثلاثي الميثَلة. لاحظ العلماء أن توزع هذه التغيرات ليس بعشوائيا, فهناك تغيرات تدل على وجود محسنات وتغيرات تدل على وجود محفزات ...إلخ. في المقالات القادمة سوف نحاول أن نشرح بالتفصيل هذه التغيرات وكيفية معالجة هذا النوع من البيانات. في هذا المقال نكتفي بالصورة التوضيحية في الأسفل.
طرق دراسة البنية الثلاثية للجينوم
طور العلماء مجموعة من الطرق لدراسة البنية التشكيلية للجينوم على صعيد المستويات الاربعة التي نوهنا لها سابقا. فلدراسة نتظيم الجينوم على المستوى الأول والثاني طورت لدراسة تموضع النكليوزومات ( nucleosome positioning) كطريقة FAIR و MNase-Seq. لكن في هذا المقال سوف نركز على التعريف بالطرق المستعملة لدراسة بنية الكروماتين على المستوى الثالث أي دراسة العروات (او الحديث المتبادل بين لفائف الكروماتين, chromatin crosstalk).
دراسة عروات الكروماتين باستعمال تقنيات السَلْسَلة
بالاضافة إلى المحفزات التي تتموضع قريبا من بداية الجين من أجل بداية تحفيز عملية النسخ يتم التحكم في التعبير الجيني عن طريق عناصر منظمة أخرى تكون في بعض الأحيان متواجد في مناطق بعيدة جدا عن الجين. يتم الاتصال بين هذه العناصر المنظمة والمحفزات الجينية عن طريق التفاف الكروماتين وبالتالي يمكن للعناصر التي تكون بعيدة خطيا (بالنسبة للوضعية في الحمض النووي) من الاتصال. 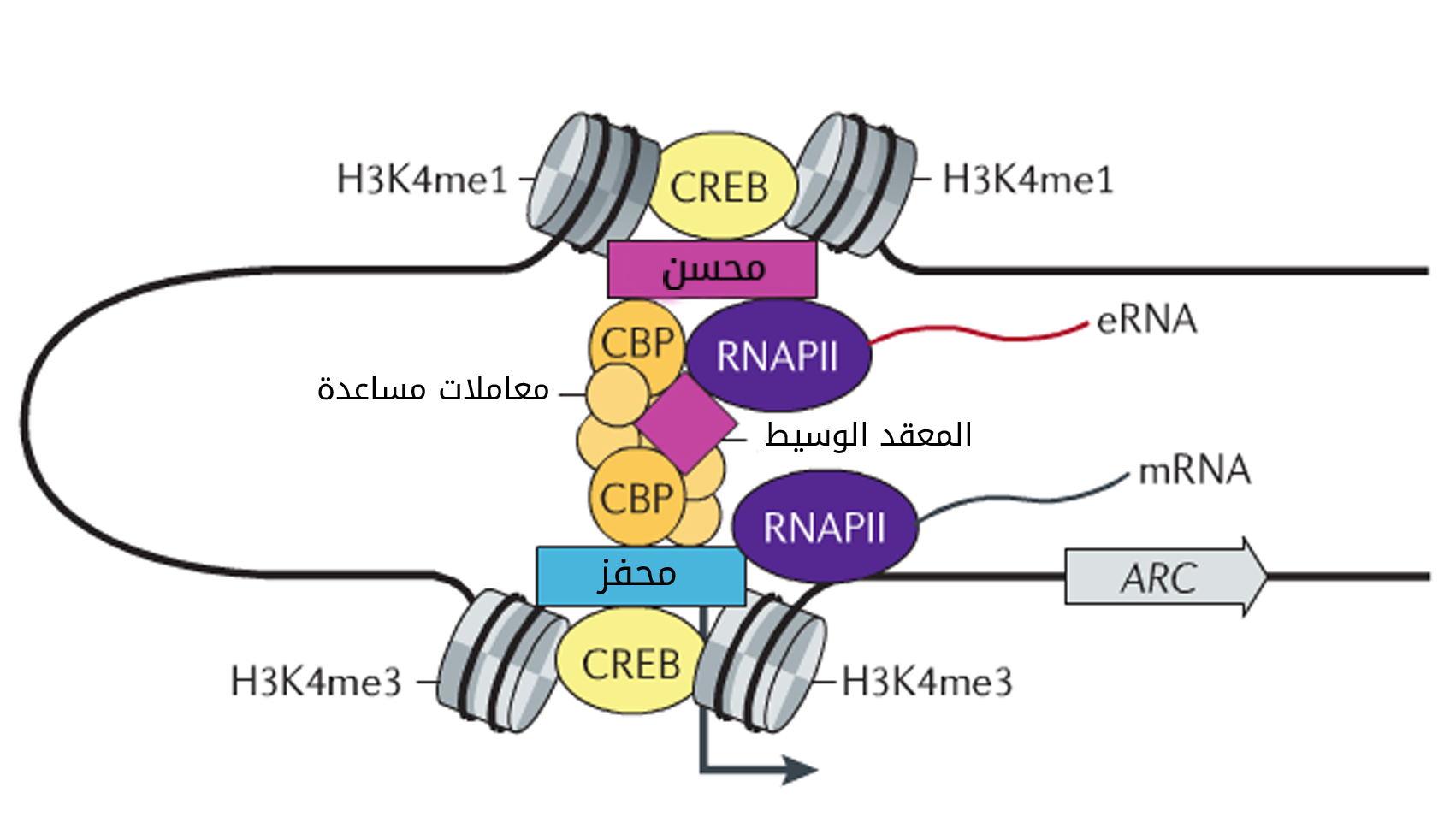
رسم تخطيطي يوضح تعديل المحسنات للتعبير الجيني عن طريق تكوين عروات (مصدر الصورة)
تم تطوير عدة طرق لدراسة الاتصال بين العناصر المنظمة والمحفزات لكن أدى استحداث طريقة التقاط التشكل البنيوي للكروماتين (Chromatin Conformation Capture) أو اختصارا 3C إلى احداث قفزة نوعية في هذا المجال وابتكرت عدت بناءا على مبدأ طريقة الـ 3C. في هذا المقال ولعدم أطالة المقال مما هو عليه سوف نحاول شرح ثلاثة طرق والمتمثلة في 3C, Chiap-PET و الـ Hi-C.
- طريقة الـ 3C:
تهدف طريقة الـ 3C إلى دراسة المناطق التي نشك أو نعلم بوجود أتصال بينها (مثلا منطقة
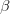 -غلوبين). تقوم هذه الطريقة باستعمال محلول الفورمالدهيد (formaldehyde) من أجل تثبيت بنية الكروماتين داخل النواة, بعدها يتم استعمال أنزيم قاطع ( restriction enzyme) من أجل تقطيع الحمض النووي إلى قطع صغيرة واستعمال أنزيمات تقوم بتلحيم قطع الحمض النووي القربية من بعضها البعض ( التي ممكن ان تشكل عروات). يتم بعدها إلغاء تثبيت الخلية و تضخيم المنطقة المراد دراستها باستعمال طريقة الـ PRC. ثم نقيس قوة الارتباط بحساب عدد النسخ.
-غلوبين). تقوم هذه الطريقة باستعمال محلول الفورمالدهيد (formaldehyde) من أجل تثبيت بنية الكروماتين داخل النواة, بعدها يتم استعمال أنزيم قاطع ( restriction enzyme) من أجل تقطيع الحمض النووي إلى قطع صغيرة واستعمال أنزيمات تقوم بتلحيم قطع الحمض النووي القربية من بعضها البعض ( التي ممكن ان تشكل عروات). يتم بعدها إلغاء تثبيت الخلية و تضخيم المنطقة المراد دراستها باستعمال طريقة الـ PRC. ثم نقيس قوة الارتباط بحساب عدد النسخ.بما أن السلاسل التي تلتحم مع بعضها البعض يمكن أن تنتج عن طريق الخطأ, نقوم باعادة التجربة لكن باستعمال سلاسل جمض نووي معزولة (في العادة cDNA ) واستعماله تنائجها من أجل تصحيح القيم المتحصل عليها في التجربة الأصلية.

رسم بياني يوضح نسبة ارتباط المنطقة الموسومة بالخط الأسود مع المناطق الأخرى باستعمال طريقة 3C, اللونين الاحمر والأزرق يرمزان إلى نسبة الارتباط في خليتين مختلفتين(مصدر الصورة)
- طريقة الـ Hi-C :
بناءا على طريقة الـ 3C تم تطوير عدة طرق مثل الـ 5C, 4C لكن كلها تعتمد على معرفة مسبقة بوجود عروات في المناطق المدروسة. طورت طريقة الـ Hi-C من أجل السماح بدراسة كل الاتفافات في الحمض النووي وبالتالي الحصول على صورة شاملة للبنية ثلاثية الأبعاد للجينوم.
تتشابه طرية الـ HiC مع الـ 3C في بعض المراحل مع بعض التعديلات, حيث يتم في البداية تثبيت الخلية باستعمال الفورمالدهيد ثم استعمال انزيمات قاطعة (في العادة HindIII أو Ncol التي تتعرف على سلسلة محددة بطول 6 جزيئات قاعدية), بعدها يتم ملئ نهايات مناطق القطع وتعليم هذه المناطق بالبيوتين. يتم بعدها تلحيم المناطق القريبة من بعضها البعض باستعمال انزيمات خاصة ثم تقطيع الحمض النووي إلى قطع أصغر (باستعمال الصوتنة مثلا) من أجل سهولة ترسيب البيوتين. في الأخير يتم الترسيب المناعي للمناطق المتصلة بالبيوتين وسلسلتها من أجل معرفة المناطق التي تشكل عروات.
رسم تخطيطي يوضح مبدأ طريقة الـ HiC ( مصدر الصورة)
عند نهاية التجربة نتحصل على مصفوفة حيث كل سطر وكل عمود فيها يمثل منطقة من الجينوم وتمثل كل خلية نسبة الاتصال بينهما. بما أن عدد قطع السلاسل ثنائية الحد (pair-end reads) التي نتحصل عليها في طريقة الـ Hi-C والتي تكون في العادة في حوالي 10 ملايين سلسة أقل بكثير من عدد مناطق قطع الانزيم (حوالي 12 مليون في الخلية الانسانية) فانه لا يمكننا الحصول على دقة كافية لمعاينة الاتصال بين اجزاء الجينوم على مستوى الجزيئات القاعدية. في العادة نحسب نسبة الاتصال بين مناطق بحجم 1Mb.
تُمَثل نسبة الاتصال بين مختلف المناطق على شكل خرائط خرائط حرارية, وبما أن هذه الخرائط متناظرة في العادة نكتفي برسم المثلث العلوي. باستعمال هذا التمثيل لاحظ الباحثون أن الجينوم مرتب على شكل نطاقات توبولوجية (topological domains) يلتف فيها الكروماتين وتكون نسبة الاتصال بين مختلف أجزائة كبيرة. يحد بين هذه النطاقات منطقة مفتوحة تحتوي على مناطق ارتباط البروتينات العازلة (insulators), وهي مجموعة بروتينات تمنع الاتصال بين المحسنات والمحفزات وتمنع انتشار الكروماتين المغاير. تتميز هذه المناطق الحدودية بنشاط جيني, كما أنها تحتوي على السلاسل المتكررة.

رسم توضيحي يبين النطاقات التبولوجية للجينوم, نلاحظ تمركز للبروتينات العازلة CTCF في المناطق الحدودية بالاظافة الى اشارة H3K4me3 التي تميز البروتينات النشطة وتمركز اشارة الكروماتين المغاير H3K27me3 داخل النطاقات (المصدر)
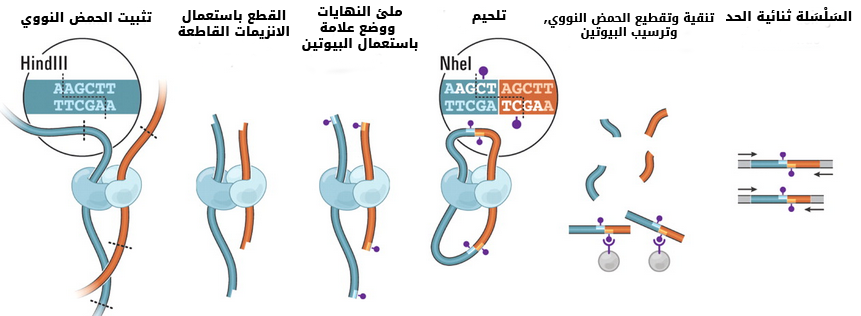
طور الباحثون أيضا بعض الخوارزميات من أجل استنباط الشكل ثلاثي الأبعاد للجينوم داخل النواة انطلاقا من الخرائط الحرارية حيث تبين أن الكروموزومات ليست موزعة عشوائيا في الخلية وانما كل منها يحتل منطقة من النواة (chromosome territory) وأن هذا التموضع يختلف باختلاف الخلية.
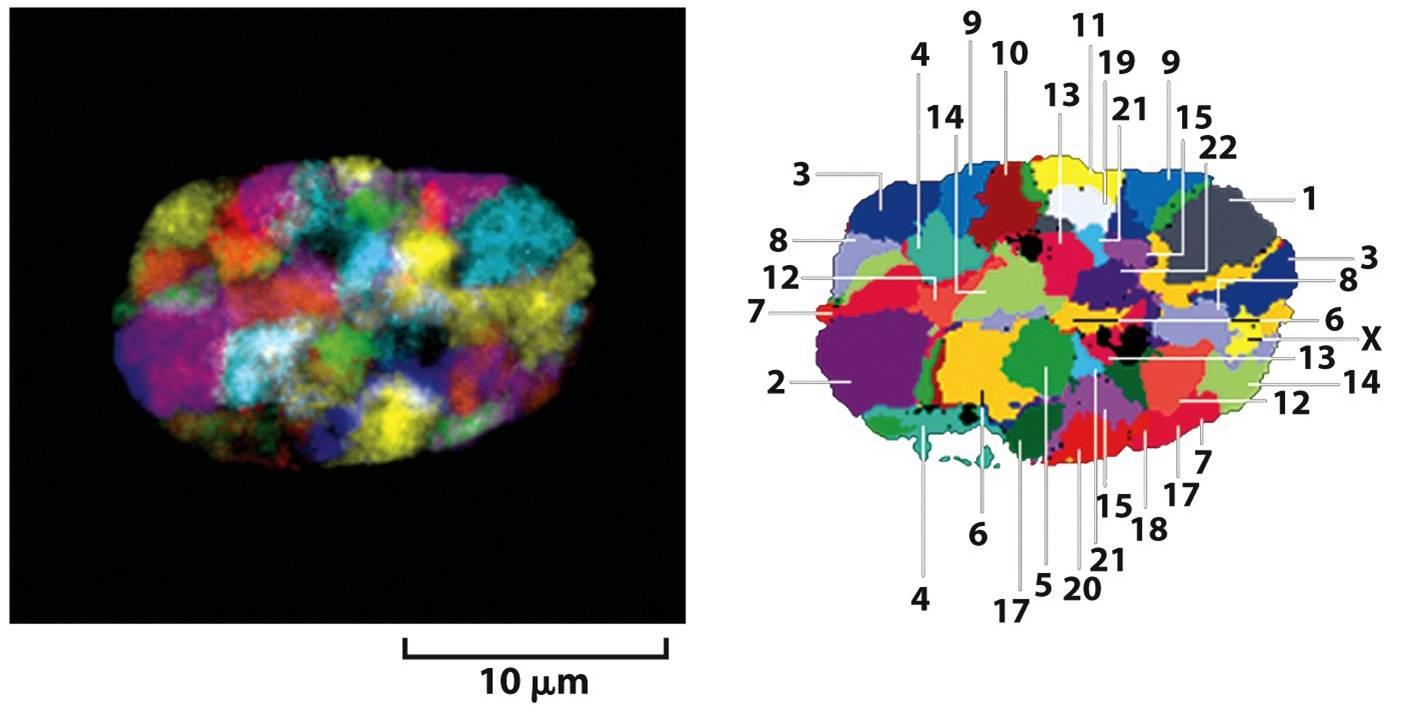
رسم تخطيطي يوضح التموضع الاعشوائي للكروموزومات داخل النواة (مصدر الصورة)
- طريقة ChIA-PET :
فيما سبق رأينا أن هناك بروتينات تلعب دورا في تشكيل العروات, مثلا بروتين البوليميراز II بالاضافة إلى بروتينات مساعدة اخرى تسمح بتشكيل عروة بين المحسنات والمحفزات. الطرق التي تطرقنا إلى سابقا لا تسمح بدراسة العروات المتعلقة ببروتين من البروتينات. لهذا الصدد تم تطوير طريقة ChIA-PET أو Chromatin Interaction Analysis by Paired-End Tag Sequencing حيث يتم الترسيب المناعي للبروتين المراد دراسته مع قطع الجمض النووي المتصلة به.
بعد تثبيت الخلية بالفورمالدهيد, يتم تقطيع الحمض النووي باستعمال الصوتنة. بعدها يتم ترسيب البروتين المراد دراسته مع قطع الكروموزوم المتصلة به, لكن قبل القيام بعملية لحم القطع القربية من بعضها البعض نظيف رابطين ( أ و ب) وذللك لتقليص نسبة التشويش حيث أنه يمكن أن ترتبط مناطق ببعضها البعض عشوائيا. بعد عملية اضافة الروابط واللحم نقوم باستعمال انزيم يقوم بقطع مسافة 20 جزيئ قاعدي قبل وبعد الرابط أ و ب. تأخذ هذه القطع ويتم سَلْسَلتها ونأخذ بعين الاعتبار فقط القطع التي الغير مندمجة (non-chrimeric أي التي تحتوي على رابطين من نفس النوع).
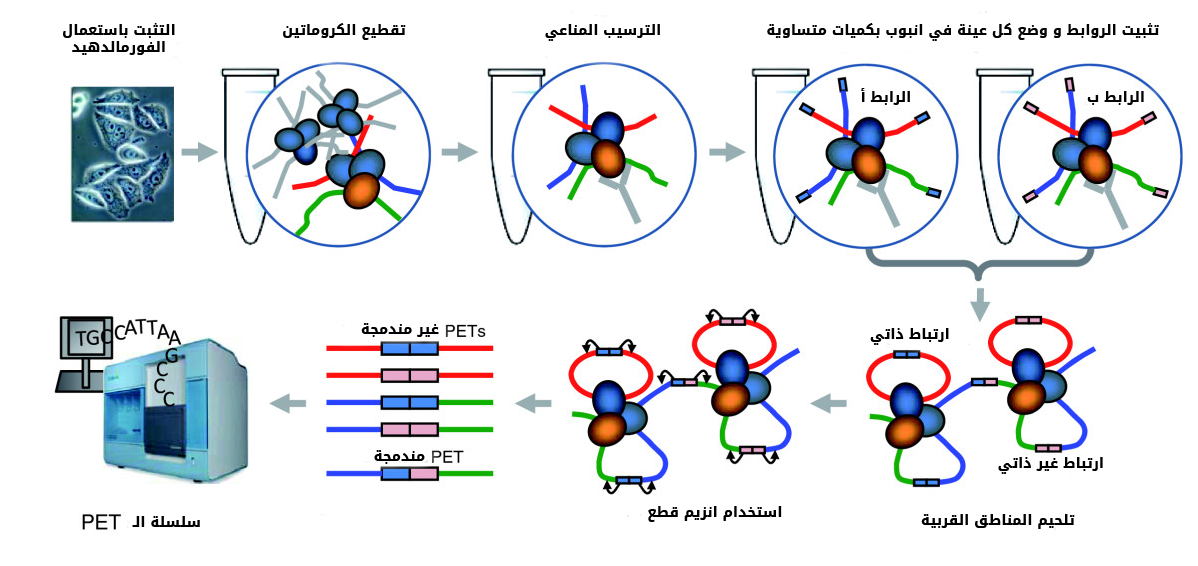
رسم توضيحي لمراحل تجربة ChIA-PET (مصدر الصورة)
لمعرفة العروات التي يشارك فيها البروتين تحت الدراسة نقوم بمطابقة الجزء الأيمن والأيسر لكل قطعة للجينوم ثم نقوم ببعض عمليات نزع التشويش الناجم عن عملية المطابقة وتحديد المناطق التي ترتبط بطريقة غير عشوائية. بما أننا نستعمل الترسيب المناعي للكروماتين فان جزء من النتائج سيكون مشابها لنتائج ChIP-Seq حيث يتطابق الجزء الأيمن والأيسر لبعض القطع في نفس المنطقة وتشكل ما يسمى بالارتباط الذاتي (Self-ligation), لاتؤخذ هذه المناطق بعين الاعتبار. أما القطع التي تتطابق أجزاءها في مع في مناطق منفصلة فيمكن أن تمثل عروة (أو اتصال) داخل نفس الكروموزوم أو تكون احدى الأطراف في كروموزوم والأخرى في كروموزوم آخر.

رسم تخطيطي يوضع أنواع العروات التي يمكن أن نتحصل عليها في طريقة ChIA-PET (مصدر الصورة)
في هذا المقال نكتفي بهذا القدر وربما في المقالات القادمة سوف نحاول ان نبرز طرق تحليل بيانات الـ Hi-C و ChIA-PET.
مصادر:
- Wit, E., & Laat, W. (2012). A decade of 3C technologies: Insights into nuclear organization. GENES & DEVELOPMENT, 11-24.
- Kalhor, R., Tjong, H., Jayathilaka, N., Alber, F., & Chen, L. (2011). Genome architectures revealed by tethered chromosome conformation capture and population-based modeling. Nature Biotechnology, 90-98.
- Li, G.et al. ChIA-PET tool for comprehensive chromatin interaction analysis with paired-end tag sequencing. Genome Biology, R22-R22.
- Lieberman-Aiden et al (2009). Comprehensive Mapping of Long-Range Interactions Reveals Folding Principles of the Human Genome. Science, 289-293.
رابط المقالة : المعلوماتية الحيوية بالعربية » البنية ثلاثية الأبعاد للجينوم